
T cell stimulation assays play an important role in modern immunology research. Not only are they useful tools in infectious disease and cancer immunotherapy studies, but they’re also critical aspects in various vaccine and drug development pipelines. Assays such as ELISpot, FluoroSpot and intracellular cytokine staining (ICS) are cornerstones of this effort, allowing researchers the opportunity to detect secreted biomarkers in vitro using patient-derived samples such as peripheral blood mononuclear cells (PBMCs).
While whole proteins, serum and live pathogens can all be used to trigger T cell activation, this is not a standardized method and is commonly associated with batch-to-batch variability. This approach can also activate other immune cell types non-specifically and induce uncoordinated cytokine release. They can introduce confounding factors into an experimental setup – such as endotoxins, immune system elements or unexpected viral/bacterial variants while also making it difficult to differentiate which epitopes are responsible for stimulation. It’s also worth noting the additional biosafety risks associated with live pathogens or serum – particularly in pipelines designed for cell therapy or drug development, where strict regulatory approval must be considered.12
Peptide pools are a game-changing technology when it comes to T cell stimulation research. While their concept and design are relatively simple, they offer superior specificity, robustness and ease of handling when compared to their live pathogen, serum or whole antigen counterparts. This article highlights the power of peptide pools in T cell research, exploring the mechanisms of T cell stimulation and the future of peptide pool applications.
While it’s easy to think of T cell stimulation as an all-or-nothing principle, it’s important to acknowledge the nuance and variability in this biological system. T cell responses (to a pathogen or antigen) are not haphazard reactions; in fact, they are highly specific, coordinated responses to small fragment regions of a foreign or stimulatory protein – not an entire organism or macromolecular structure. These small fragment regions, known as epitopes, are recognized through complementary binding to T cell receptors (TCR) on lymphocyte (T cell) surfaces. For a T cell to be activated, its TCRs must bind to specific epitopes and initiate a downstream intracellular signaling cascade. As TCRs themselves possess no inherent enzymatic activity, activation is mediated through protein kinases and ultimately transcription factor activity.3
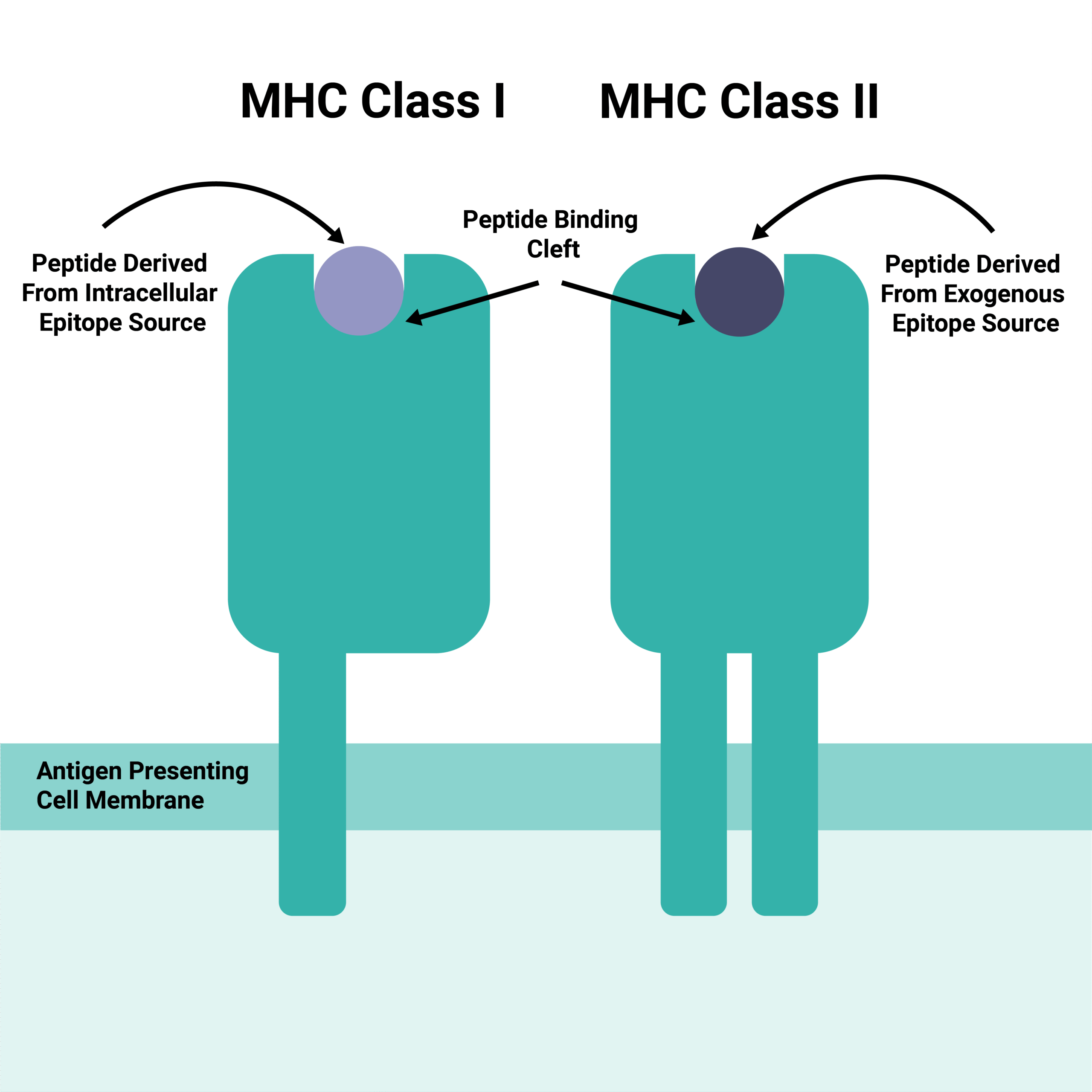
TCRs recognize epitopes derived from both intracellular and extracellular pathogen proteins. Crucially, they don’t bind to the raw epitope sequence; instead, they bind to processed molecules presented on the surface of antigen-presenting cells (APCs).4 These APCs feature Major Histocompatibility Complex (MHC) molecules, specifically Class I and Class II. APCs first internalize and process the viral and bacterial epitopes before displaying them within the MHC binding cleft.5 Intracellular epitopes (like those from nuclear or cytosolic proteins) are typically presented on MHC Class I. Conversely, extracellular epitopes (such as those from capsid or vesicular proteins) usually present on MHC Class II, although there are through to be some limited exceptions. Both MHC Class I and MHC Class II presentations stimulate different T cell subtypes (CD8+ and CD4+ respectively).
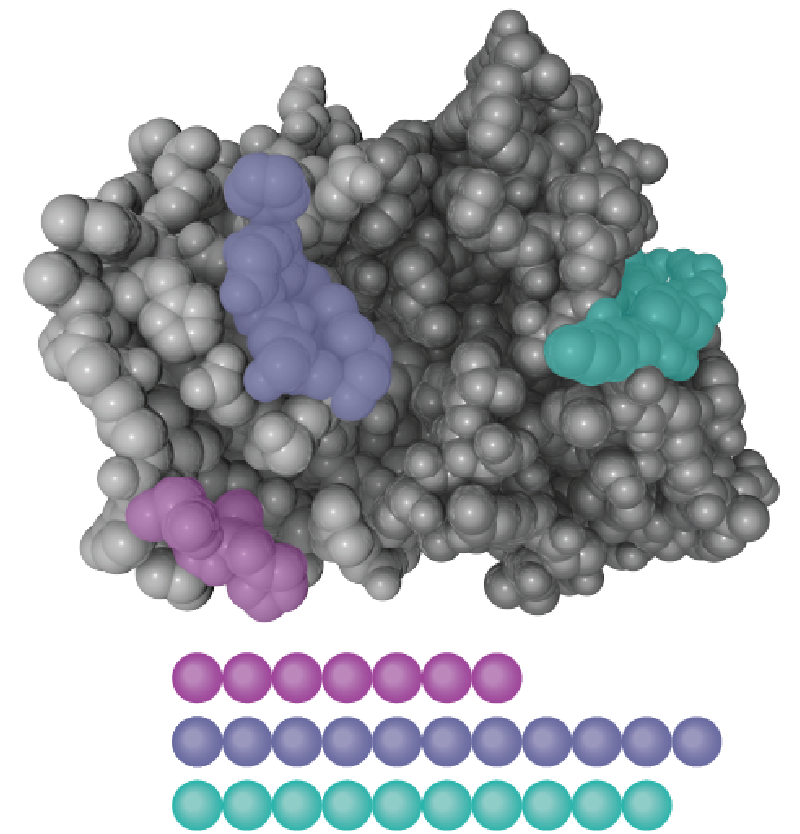
Not all epitopes are equally effective at eliciting a T cell response. In fact, all epitopes exhibit varying degrees of stimulatory signaling, also known as immunodominance. While the exact mechanisms underpinning immunodominance are still debated, some factors may include the relative expression (concentration) of the epitope or genetic/protein-level variant expression in the pathogen or host.6 Identifying and using only the most immunodominant epitopes can be one way to strengthen and control T cell stimulation assays and remove confounding factors. Furthermore, it can help researchers better understand the factors driving an immune response, define precise antigenic targets, or evaluate novel therapeutics. Many immunodominant structures are already reported in open-source databases such as the Immune Epitope Database (IEDB).
Peptide pools offer a strategic solution to the challenge of epitope and immunodominance variability. They can be precisely engineered in the lab using biochemical methods to collectively reflect the major immunodominant regions of an antigen or pathogen of interest. They may be engineered as overlapping or non-overlapping sequence constructs, depending on the need for structural coverage.
While overlapping peptide pools are important for epitope mapping, non-overlapping sequences are ideal for ensuring the robustness of immune stimulation; they allow researchers to target multiple immunodominant regions, optimizing T cell stimulation in response to known TCR/epitope sequence pairings.
Unlike whole proteins (which must undergo complex and sometimes inefficient intracellular processing), peptide pools also bypass a major bottleneck in the immune signaling pathway. Being relatively small, they’re ready to be internalized and shown on the surface of antigen-presenting cells (APCs) without the need for additional, biologically variable cleavage steps. The resulting, highly controlled presentation of immunodominant epitopes further ensures comprehensive coverage, superior stimulation and more reliable data. Through clever, expertise-informed design, peptide pools can also be engineered with MHC Class and immune cell subtypes in mind; in fact, products can be designed to cover both Class I and II with equally potent immunogenicity, providing an in vitro model that accurately reflects infection biology without introducing undue variable factors.
The advent of synthetic peptide pools for immunology research is enabling scientists and clinicians to conduct far more accurate and scientifically valid analyses of T cell stimulation than ever before. By moving beyond the limitations of whole antigen, serum and live pathogen stimulation options, it is now possible to achieve comprehensive epitope representation without compromising experimental design and while maximizing workflow biosafety. Peptide pools not only overcome the challenges of unknown variant factors and genetic polymorphisms but also enhance the sensitivity and specificity of immunological assays. As a result, peptide pools have become an indispensable tool for immunologists, providing a powerful and reliable method to explore cellular immune responses with unprecedented precision.
**